
This year was a mast year of conference-going for me. I hadn’t planned to attend nine conferences (summary of each at the end of the post), but sometimes it is so hard to say no! However, I will not be repeating this schedule in the future, as it has been very hard to keep on top of all the other work I want to/ have to do. So how should I decide on which conferences to attend in the future?
I think the basis for the decision will differ from year to year depending on what I hope to gain, whether I have a new exciting story to present, and whether I am hoping to be inspired and think outside of my usual scientific framework, or hoping to network and build on what I am currently doing. But generally I think meetings break down into three categories:
(A) the large society meetings that feel more like a family reunion than a day at the office;
(B) the specialty meetings where like minds come together and reinforce the momentum of the field, and;
(C) the new topic meetings – either altogether new, or simply new to me – where I spend the entire time brainstorming about what direction I want to move in next and networking is the name of the game.
Each type of meeting has its pros and cons, and I hope to always balance among the three (which means missing the “family reunion” from time to time). Type A acts as a reminder of why I am doing what I do, how much I enjoy my colleagues’ and advisors’ company, and reinforces that I have a terrific support network. These meetings typically have an overwhelming number of talks you could attend (transports me back to the excitement of opening my University course offerings directory each term), are less restrictive in who can give a talk – allowing for early career researchers to present their work, and are highly social. Type B is where I typically gain the most in terms of scientific progress and networking. These meetings often don’t have concurrent sessions, which means that everyone has seen the same set of talks as you, and therefore that dinner conversation topics are easy, even with folks you’ve never met before. This also means you are likely to sit through a talk you wouldn’t have chosen from an abstract book, and learn something unexpectedly relevant to your own interests. Type C are also terrific for networking, as they are often very small and targeted, with a particular goal in mind (synthesizing a field, solving a specific problem, or building collaboration). These meetings are probably the most likely to result in the generation of a new project, and can really solidify a community of researchers. So if you see one on a topic you are interested in getting involved in, there is no better way to put your name on the map than attending!
As you can see, there is no easy choice (especially if you are resource- and/or time- limited). One way to make the decision, especially if you are further on in your career, is to only attend the meetings to which you are invited, either as a plenary speaker or within a symposium. However, if you follow that track you may miss new opportunities for networking, as you are unlikely to be invited to a meeting that is outside your area of expertise. My new plan is to choose one meeting that I want to attend, one meeting that I “should” attend (i.e. based on the table below), and then one that I am invited to, unless that fills either of the two previous niches. I’ll let you know how that pans out next season.
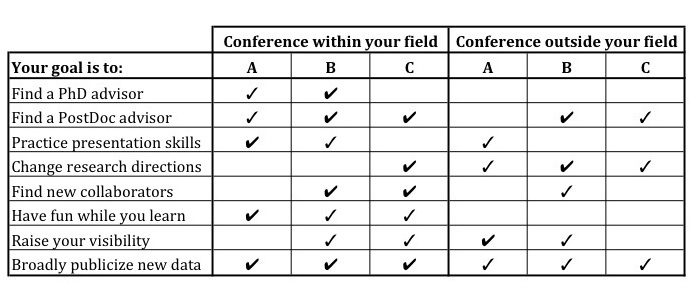
A bit about this year’s conferences:
First I attended a meeting in Eco-Evolutionary dynamics in Leuven, Belgium which was organized by Ellen Decaestecker and Luc De Meester. The meeting was primarily designed as a networking event for Belgian researchers, but I think all of us invited speakers found it just as useful for networking! The best thing about this meeting was that although it was specific in terms of question being addressed, it was incredibly broad in terms of systems and approaches being used. It was also a great group, with the bulk of the audience representing PhD students. The questions after each talk were very insightful, and it was a warm open atmosphere. The research highlights were hearing about hell breaking loose in three species lab experiments from Nelson Hairston, data presented by Lutz Becks showing increased sexual reproduction with ecological feedbacks, experimental work by Eva Lundstrom indicating that the abiotic environment can select for particular bacterial communities even in the face of 70% dispersal, and a fabulous talk by Jacintha Ellers about trait loss in obligate parasites.
Next I went to the Second International Microbotryum meeting in Amherst, Massachusetts. The meeting was organized by Michael Hood, and focused primarily on using the Silene-Microbotryum system as a model for understanding disease ecology and evolution. However, since many of us have changed paths since studying that system, it included talks from across many plant disease systems. Due to the size of the meeting, everyone presented their work, including some terrific undergraduate students, which meant that we all really gelled and could have great/productive conversation. I had never met Pete Thrall, Tatiana Giraud (although we’ve written a paper together) or Georgiana May, and really enjoyed getting to know each of them. It was an ideal size for a meeting of this sort (20ish), and is even leading to a new publication. Among the highlights and new things I learned were: Emme Bruns explained the use of Aster models for life history (I am definitely going to use these!), Janis Antonovics showed both theory and data on disease spread at the edge of host ranges, Scott McArt taught me about mummy berry disease (very cool!), Britta Buker shared a really great way to identify genes important in host specificity by backcrossing hybridized parasites, Martin Kemler described successful Microbotryum infection of non-host species but no onwards transmission, and Mandy Gibson showed great support for the idea that sexual reproduction is higher in parasitic than free living species of nematodes.
The month of May was a great one for folks studying the evolution and/or ecology of infectious diseases. First, we got to attend a one-day symposium in London organized by Mike Boots on the Evolutionary Ecology of Infectious Diseases (as part of the British Ecological Society’s 100th birthday!) Then we flew over to Pennsylvania State University for the 10th annual Ecology and Evolution of Infectious Disease meeting (and Pete Hudson’s 60th birthday celebration). Both meetings were fabulous! And it became increasingly clear that our understanding of when/how/why disease occurs is dramatically improving. For one thing, there are more researchers working on these questions than ever before; but also, there are some exciting new techniques and approaches being applied, both in generating empirical data and in modeling large datasets. It seems to me that the EEID field has long been lead by theoreticians, with very little data that could be used to test the elegant and clear theory being produced. The new datasets being generated and modeled are pushing the field forward and challenging many of the ideas we thought we had figured out. And this is really exciting – new data leads to new theory leads to leaps and bounds in our ability to manage disease.
The research highlights of these two meetings were much too plentiful to put into this post, but I did live tweet both events, and have collated those here: Tweets from EEIDs. The annual EEID meeting is among my favorites on account of its smallish size, great social events, fabulous and diverse talks, and lively discussions at the end of each session. This time around, the moderators of each session were in charge of shaping the timings for discussion and really got things going by asking provocative and insightful questions. For more highlights from the meetings see: http://monkeysuncle.stanford.edu/?p=1201 and http://parasiteecology.wordpress.com/2013/05/24/parasiteish-host-coevolution/
Next up was Evolution 2013 in Snowbird Utah. Unfortunately, I missed the opening night and first day, and it took me some time to recover from jet lag and altitude once I arrived. Despite these obstacles, however, it was great fun as always. I was lucky enough to be part of the ASN Vice Presidential symposium on Ecology, Evolution and coevolution of host-parasite interactions, organized by Curt Lively. It was a great experience, and a very supportive group to be part of. Topics ranged from the role parasites play in maintaining sexual reproduction (Curt Lively), bacteriophage adaptation over space and time (myself), self-medicating butterflies (Jaap de Roode), parasite-mediated competition (Meghan Duffy), eco-evolutionary feedbacks in immunity (Andrea Graham), to parasite evolution in immune-compromised hosts (Andrew Read). In the next year or so, you’ll be able to read all about the work presented in a special issue of AmNat.
As always, the meeting was jam-packed with sessions. Unfortunately, it was quite difficult to move among rooms with ease, so I tried to stay through whole sessions. In many ways I prefer this to running around madly, but it did mean I missed many talks I wanted to see. This year’s Evolution meeting also had a new twist: Lightning talks. These 5 minute shorts were a great test of whether a researcher really understood what their work was all about. The best ones typically had only one key point being made; some were funny, others were truly lightning paced, but all were good effort! Overall, the meeting was in a great location (although I didn’t have time to fully exploit it) and was a wonderful opportunity to see many many friendly faces. Again, for more coverage see: http://roadsidescience.com/2013/07/17/host-parasite-interactions-highlights-from-evolution-2013/
After a three-week break it was on to the Gordon Research Conference on Microbial Population Biology in New Hampshire, organized by Paul Turner. This is my second GRC, and I am sure it won’t be my last. The success of the GRC meetings rests on the quality of the talks and posters (which are almost exclusively on unpublished results), the isolated location of the conference (think summer camp), and the fact that it draws the big names from the field (i.e. great networking opportunities!) Given the confidential nature of the talks – no live tweeting allowed – I will only give vague research highlights here. There were two talks about reconstructing ancestral phenotypes, one by Betul Kacar and one by Eric Gaucher, which really got my imagination going, one about megaplasmids in Pseudomonas syringae by Dave Baltrus, a full session on CRISPRs, including a glimpse into how spacer heterogeneity evolves by Rachel Whitaker and a discussion of the non-random acquisition of protospacers by Rudolpho Barrangou, and an amazing illustration of the evolution of resistance to antibiotics in a Guinness Book of World records-sized agar plate by Roy Kishony.
For those planning to attend the GRC for the first time in coming years, one thing to note: although the meeting can seem “cliquish” at first glance, I’ve found that the meal layout with round tables after a buffet choice is perfect for meeting new people. Whatever you do, don’t stand there looking around for the one person you know. Go to the nearest free seat and ask if you can join. In fact, this should go for every meeting you attend, but it is easier said than done at some.
That brings us up to date, with a nice two-week respite before I head off to the European Society for Evolutionary Biology meeting (ESEB 2013) in Lisbon, Portugal, followed by a desperately needed 10 day holiday in Italy, followed by the Society of General Microbiology meeting in Brighton, UK, followed by two talks (one on science and one public lecture on “what evolution can do for you”) at the 2013 EMPSEB (European Meeting of PhD Students in Evolutionary Biology) meeting organized by our very own PhD students here in Cornwall.
As I said, I won’t be repeating this grueling schedule anytime soon. But it’s been great fun so far!

Oops, make that 10! I nearly forgot the BSPP “Protecting our woodlands: Tackling tree pests and diseases” meeting in September in Reading. That should be great fun, and hopefully very productive too.
Interesting Read